

渡り鳥は帰る場所を持つから、遠くへ行けるのよね。ケラトプスユウタです。
2025年、我孫子市鳥の博物館で特別展やってたんですよ。『とりホネ展-骨からわかる鳥の進化と多様性-』という恐竜展。
我孫子市鳥の博物館は日本最古(1990年開館)の恐竜専門の博物館にして日本初の鳥類専門の博物館でもあります。
と言うわけで早速レポートと称して標本の写真を紹介します。


飛翔に利用される骨や筋肉、腱の構造がわかりやすく標本一体で紹介されています。翼の羽と尾羽も残されています。
翼の羽のうち、初列および次列風切は羽ばたきのエネルギーを体に伝えるため、骨にしっかり付着しています。尺骨の羽の付着部をチクビに例えて尺骨乳頭 ulnar papillae とも言います。僕は標本を見た事ないのですが、ヴェロキラプトル Velociraptor のような非鳥類型マニラプトル類 Non-Avian Maniraptora でも報告されています(Turner et al. 2007)。

鳥以外の鳥頸類 Ornithodira 等やワニ以外の偽鰐類 Pseudosuchia が絶滅している関係で、現生種だけを見た場合、ワニ類 Crocodylomorpha が鳥類の姉妹群に当たるのです。
他にも哺乳類やカメなどの骨格も鳥類との比較のために展示されていました。

ハクスリー(キャプションではハックスレー) Huxley による頭骨の口蓋面による鳥類の分類の解説がありました。
シギダチョウは唯一飛翔能力を残している古顎類として知られています。

腸骨の下に仙椎だけでなく胴椎や尾椎もかなりの個数隠れているのが観察できます。ほぼ腸骨の前後長=大腿筋の前後長らしいので、腿は短いながら逞しい筋肉がついていたことが想像できます。この写真ではわかりにくいですが。

哺乳類の頸椎は一部のナマケモノ類を除いて基本的に7個ですが、オウム類は10個もあります。それでも鳥類では最少の個数です。巨大な頭を支えるために短い方が好都合という点でケラトプス類 Ceratopsidae やティランノサウルス Tyrannosaurus に通ずるものがあります。

ハクチョウ類の頸椎は26個で鳥類最多です。これは僕の考えでは体を手入れしたり摂餌したりする為だけでなく、鳴き声の遠達性を高めるための進化の一環もあるのではないかと思います。

第七頸椎が短く、首を縮めて勢いよく伸ばすのに役立つらしいです。

飛翔して獲物を捉える姿勢。この展示では飛翔能力と肩帯の構造の関係について説明されています。

鳥やそれに近い恐竜類が翼をおりたたむと、前肢はZ字状になります。知ってますよね。

鳥類とコウモリ類の翼の構造の違いが解説されていました。

ワシミミズクの目はほぼ管状で、その形を支えるのが強膜輪です。ご存知の方は多いと思いますが、強膜輪は複数の小さな板状骨 ossicles がタイル状に連結して環を作る構造で、化石でも保存されることがあります。
ワシミミズクの強膜輪は暗所視に特化した望遠レンズ設計です。強膜輪の直径が大きいほど、より大きな瞳孔を確保=わずかな光源からより多くの光を取り込む事ができます。ワシミミズクの強膜輪は夜行性鳥類の中でも最大級らしいです。

基盤的なものはどうか知りませんが、鳥類は概して左右の腸骨の挟み込みがめちゃくちゃ高度に発達して仙椎を完全に覆い隠しています。この部分の発達がすごいと言われるティランノサウルスでも明らかに腸骨間の間隙はあるので、かわいく見えちゃいますな。

生体や剥製のクチバシには角質のセレーションのような細かい突起があり歯にような役割をもっているのですが、クチバシの骨は滑らかで痕跡すらありません。化石恐竜でも下クチバシの角質の鞘の縁がギザギザしたものが知られていますが、意外な恐竜がこのような構造を持っていてたとしても骨だけではわからないですね。

Recurvirostra avosetta
その名の通りそり返ったクチバシがユニークです。

脛足根骨(すね)と足根中足骨(足の甲)の長さを見てください。この足でヘビを蹴るんですね。

かわいいです。
キツツキのなかまは全てかどうかはわからないですが、第四趾が後ろを向きます。これを外対趾足というそうで、垂直の木の幹にとまる際に役立つようです。

鼻孔がかなり切れ込んでいる様子はトリケラトプス Triceratops の巨大な外鼻孔を彷彿とさせます。海水を飲むカモメ類は、塩類腺という器官が発達しており、この鼻孔を通じて塩を排出する役割をもっているらしいです。

クチバシがメリハリのある形でかっこいいです。


カッコウとヨタカは胸骨が末広がりな形をしており伝統的には近縁な位置に置かれる事もあるそうですが、分子系統解析ではそれほど近縁ではないという事になっており、胸骨が似ているのは収斂進化によるものらしいです。どのような要因で胸骨が末広がりになるのでしょうか?

長野県戸隠に旅行で行った際、本種が木の幹に逆さま(頭を下)にしてとまっているのを頻繁に見ました。彼らが木の幹に逆さにとまれるのは、後趾(第4趾)が特に発達しており、鋭く強く湾曲した爪が樹皮に食い込むことによって、足だけで体を支えられるためらしいです。尾で支えなくても保持できる点が、他の多くの鳥との大きな違いとされています。

川上先生の一般書『鳥の骨格標本図鑑』によると、夏の方がクチバシが長い傾向があり、夏冬で1mmも長さが変わることがあるそうです。

尾羽が短いために日本最小級の鳥となっている本種ですが、やっぱし尾椎もそれなりの短さでした。

離陸直後の姿勢。
さすがは渡り鳥で、竜骨突起は前後に長いですね。

カモと比べて前肢がとても長く、肘が大腿骨の付け根よりも後ろに位置しています。
体重は体長の3乗で増す一方、翼面積は二乗でしか増えないため、体が大きくなると体重を支えるために翼のサイズを相対的に大きくしなくてはならないそうです。何言ってるかわからない人は読み飛ばしてください。

シジュウカラガンと比べると前肢の短さがわかります。

有羊膜類 Amniote の気管は喉から下に一直線に向かい、肺の手前で二又にわかれるのが普通です。
しかしツルのなかまでは分岐する前に気管が胸骨の中に入ります。そこで複雑に折り返し、最終的に胴体内に向けて出てきているらしいです。
キャプションによると、長く伸びた気管が胸骨を震わせ、胴体を共鳴器としているという事のようです。
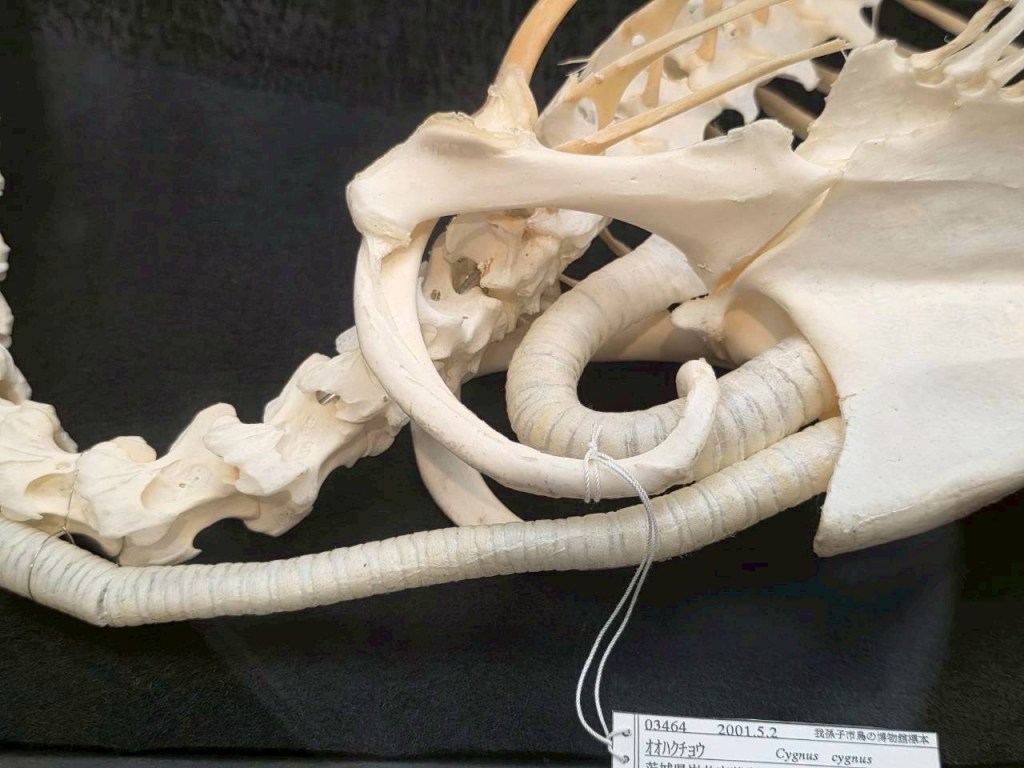
気管が胸骨の中に深く侵入し、一旦ループ状に折り返して肺へと繋がっているらしいです。チューバのように、音声は管が長いほど低くなりますが、首の長さには限界があります。そこで胸骨内部に気管を巻き込むことで長さを延長しているようです。それだけ声が遠くに響く方がモテるという事なんでしょうね。
ハクチョウとツルは鳥類の中では近縁ではないので、どちらも気管を胸骨に侵入させ、管共鳴型の原理で低い声を出すという進化を独自に遂げた事になるんですが、これは並行進化で説明していいのでしょうか?

本種もラッパのように啼哭するらしいですが、案の定胸骨に気管を入り込ませている口でした。

本種は多くのカモが地上に営巣する中、樹洞に営巣したり、カモなのにドングリをよく食べるという変わった生態ですが、そういった生態は一見して骨格には現れていないようで、骨格はいたって普通のカモのようです。骨だけで化石恐竜の生態を論じる事の限界を感じますね。

キジの雄は足根中足骨からケヅメと呼ばれるトゲが生えているのが特徴です。これでオス同士やり合おうという魂胆です。蹴爪と書きますが爪ではなく、ヒトで言うと踵からトゲが出ているようなものです。皆さんはこれ、自分の踵に欲しいですか? 僕は腿裏とか足の甲側だったら欲しいです。

人体模型みたいに右半分が骨格標本で左半分が剥製というすごい職人技のひかる標本。

眼窩の上に眉のように骨が飛び出ているのがわかりますでしょうか。この骨は多くのタカ類に見られる構造です。ジェームズ・キャメロンの映画『アバター』のトルークと同じで、頂点捕食者は上空を警戒する必要はありません。上方からの余計な光をカットするカメラのレンズフードのようなものらしいです。この理屈は非鳥類型獣脚類にもよくある涙骨だの頭頂骨だの由来の突起とも関係あるかもしれませんね?


これも翼の羽と尾羽を残した標本ですね。

島間移動や海上飛翔が必要な生活史のため、竜骨突起が深い=胸筋の付着面積が大きいらしいです。森のハトというより海を越えるハトの体つきだそうです。上のキジバトの画像とこうやって交互に比較してみると…正直微妙です。なおカラスバトは系統的にはキジバトよりも、同属別種のドバト(カワラバト) C. livia の方が近縁なのでそっちの方が比較には向いているはずです。

ツグミ類 Turdidae は典型的な止まり木型鳥類で、跗蹠骨が比較的長く、趾骨の長さの比率が枝を握るのに最適化、後趾が強く反転といった特徴は、地上歩行中心のハト類等とは明らかに異なるつくりらしいです。

初めて知ったのですが、カワセミ科は、生体では第3趾と第4趾がくっついた合趾足という状態になっています。ところが骨だけにすると2本の趾が癒合せずに普通に存在しています。これは地中や樹洞で土や木片をかき出すのに使われるそうな。

足根中足骨が異様に短いのがおわかりいただけますでしょうか。つまり脛のすぐ後に足指が伸びている感じです。この短足は進化の証であり、枝上からのわずかな助飛距離で体ごと水中に突き刺さる上で、体から外側に飛び出てしまう足がなるべくコンパクトになるという自然選択の帰結らしいです。

カワセミ以上の短足(足根中足骨が短い)です。川上和人先生の言葉を借りれば、ヤマセミの狩りの様子は「サギが頭だけで飛んでいるよう」。

3色のクチバシがチャーミングな本種ですが、角質の鞘は骨と同じ形をしていませんね。骨だけだと、ちょっとグリポサウルス Gryposaurus のような鼻骨が隆起した鳥脚類 Ornithopod を彷彿とさせます。

しゃもじのような上下のクチバシをこうやってよーくみると👇

立派な血管神経孔がたくさん空いているのが見えるではありませんか!
クチバシに多くの感覚器官が配置されている鳥はみんなこうなっているそうです。我孫子市鳥の博物館の湧水先生や福井県立大学の河辺先生らは中生代恐竜等のこの部分の研究の権威です。少なくとも僕はそう思っています。

一部の大型種を除き、サイチョウ類のカスクは基本的に種内闘争用の打撃武器ではなく、共鳴器らしいです。カスク内部は空洞化しており、頭骨の気嚢系と連続しているそうです。これにより、鳴き声の音響増幅が起こるとか(Alexander et al. 1994)。
森林環境で遠くまで声を届ける工夫(工夫ではないですが)なのでしょう。

長いクチバシを砂浜にぶっ刺して甲殻類を捕食するそうです。容易に目に浮かびますね。

いやはやいやはや。強靭な叉骨をお持ちですな。
これはなかなか鳥以外の恐竜では見られないつくり。鳥盤類はそもそも叉骨がありません。
羽ばたきのたびに肩帯は外側へ引き離されます。このとき叉骨はわずかに開き、戻ります。もしまっすぐだったら応力が一点に集中し壊れやすく、弾性エネルギーをためにくいですが、湾曲していると応力が全体に分散、骨が元の形に戻ろうとする力を効率的に生むことができると説明されます。弓と同じ原理ですね。

全長の半分ほどが尾であるオナガも、長いのは尾羽だけで尾椎は短いです。現生鳥類で翼の爪や歯っぽいものなど爬虫類じみた器官を再発達させるものはいますが、どんなに尾が長い鳥も尾椎を伸ばすようなバカなマネはしてないみたいですね。

周日行性の為、眼窩が昼行性の他のツグミ類より大きいらしいです。

カラス科 Corvidae はスズメ目の中では体が大きい事とクチバシが大きい事が特徴らしいです。

この標本も半分骨格で半分剥製です。
首や肩周りなんかは生体では太く見えますが、骨が細くてほとんど軟組織ということがわかります。逆に足首から先はあまり軟組織がなく文字通り骨と皮だけですね。

同じカラスでも、主に(自然界では)屍肉食のハシブトガラスと比べてクチバシがまっすぐでより採集者タイプの形質をもっているように思います。調べたところ、実際に地面をつついて昆虫やミミズ、ドングリなどを漁るらしいです。

キービジュアルやグッズを見る限り、本展の目玉展示らしいです。生体で印象的な巨大な頭部は骨でもそのままでした。

この展示が一番印象的で、個人的に獣脚類の中では一番気になる存在です。

科博の鳥展ではキャプションには書かれていたのですが、剥製では確認できなかった部分。尺骨の近位前側に「サメの歯のような形のケヅメの的な突起」と「関節つきの指のような構造」がついており、これらを使って種内闘争を行うと言うのです! どちらも本来の指とは関係ない事は明らかなので余計に面白い!
それだけでなく、飛翔能力があるにも関わらず、鉤状突起がないのです!

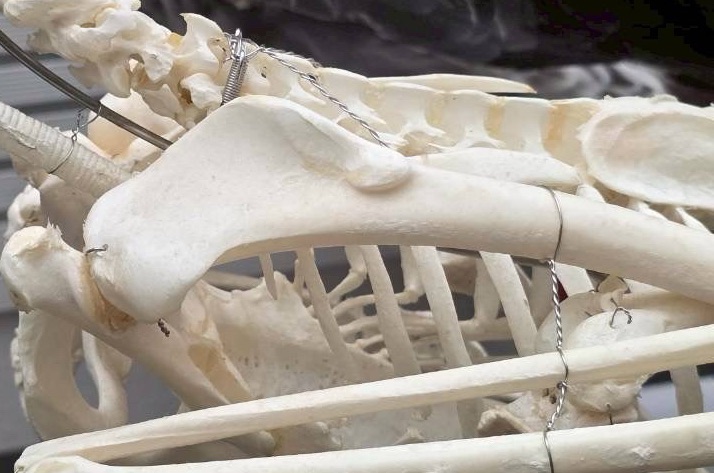
一部上腕骨で隠れていて見えないので、これは見たことがない方は実物をどこかで見ていただきたいのですが、本当に鉤状突起が影も形もなく普通の非鳥類型恐竜のようにシンプルな見た目になっているのです!
鉤状突起は何らかの形で飛翔に役立っているとされ、現生鳥類の共有派生形質であり、飛翔能力はないダチョウでも退化傾向ですが残っています。キウイにも残っています(キウイは卵がでかい関係でむしろ発達。こっちはこっちで面白すぎる)。その上、非鳥類型マニラプトラであるヴェロキラプトルからも報告されています。エミューの鉤状突起も完全に退化しているのですが、彼らは飛べないからわかるじゃないですか。

退化って、無い方が生存に有利(言い換えれば有る方が生存上不利)な場合に起こり、有っても無くてもどっちでも良い(役割はないが、有っても生存上不利にはならない)場合は起こらないものだと理解しているのですが、サケビドリの生態であれば鉤状突起は有った方が有利な場合はあれど、無い方が有利な場合というのがどうしても思いつかないので興味が尽きません。鉤状突起にまわす栄養を節約しているという安易な説明もできなくはないと思いますが、やはり飛翔能力がある事と「飛ばない歴」の長いダチョウでは消失していない事が引っかかるので説得力が皆無です。ぜひ有識者のみなさんご意見をお聞かせください。

舌骨の発達が凄すぎて末端が頭頂部に位置しています。

プテラノドン・ロンギケプス Pteranodon longiceps の後頭部から後ろに伸びたクレストのようなフォルムから展開して扇状のトサカになるユニークな冠羽をもつヤツガシラ。僕が学生時代、鳥見を始めたきっかけの鳥という話は以前しましたっけ?まあいいや。
なんと、この特徴的な冠羽の基部には何の特徴もありません。それはそれで面白いです。つまり何の変哲もない普通の頭蓋をもった化石恐竜の頭のてっぺんに冠羽をつけても許されるという事なんですよ。

口を開けながら飛び、虫を捕まえて食べるそうです。口角がだいぶ後方まで裂けているのはその為なんでしょう多分。その点は比較的遠縁のヨタカと似た様子だと思います。

太短いクチバシで水面の種子や葉を食べるそうですが、「太短い」ってたいてい強暴な力にさらされる器官の構造じゃないですか。本官、「ほんとうにそれだけか?」と疑ってしまいます。


アビ類やカイツブリ類は後肢が体のけっこう後ろの方についてるらしいんですよ。でもその脚の位置が、水の中だとすごく都合がいいらしく、後ろからしっかり水を蹴れるので、いわばプロペラかスクリューか何かが潜水艦のちょうどいい場所についてる感じだそうです。
後肢が後ろすぎてバランスの問題で地上では歩けない(這うような動きになる)とよく聞くんですけど、本当ですかね?
翼は飛翔と舵取りに使い、水中での推進装置としては使わないようです。

水面性のカモは、オオハムのような潜水特化型の鳥ほどではないですが、脚がやや後方寄りについているようです。遊泳と歩行の折衷案でしょうか?

サカナじゃない方のウミスズメ。オオハムやカイツブリの翼は完全に飛翔特化で水中では使われないのですが、ウミスズメの翼は真の水空両用で飛翔と遊泳の両方に使われるそうです。かっこいい!

体は小さいですが、同サイズの鳥と比べて相対的に足ががっしりして見えます。押さえつけて鋭いクチバシで肉を食いちぎるつもりに違いありません。恐い!


獰猛凶暴なモズは江戸時代には猛禽類に含められていたと聞いたことがあります。
赤髪のシャンクス「どのみち(猛禽類)は多系統群だろう」
まるでハヤブサのような歯状突起 tooth が上クチバシにあることからもその性質が窺い知れようというもの…
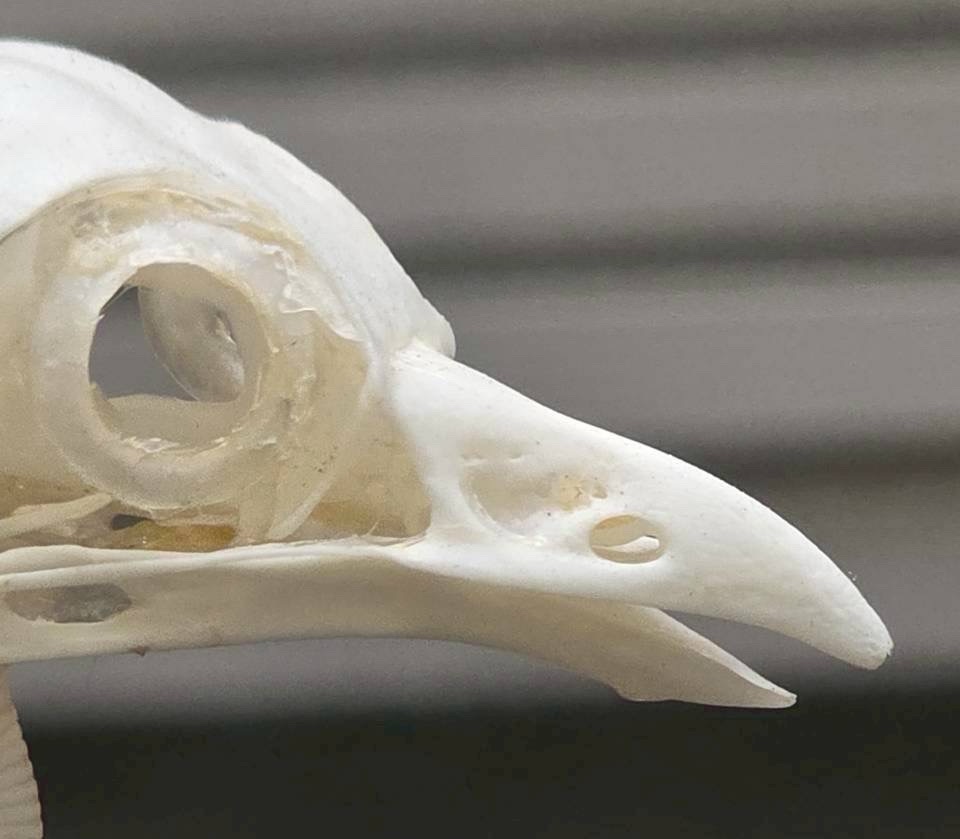
…しかしながらクチバシが曲がっているのは角質の鞘だけで骨は直線的で大人しい形をしています。はじめて知りました。角質の自在さも知りました。

盗賊のイメージとは裏腹に、クレプトパラジティズムの頻度は少なく、自力でサカナやイカを捕える事が多いそう。まあ少なかろうが多かろうが、一度やったら窃盗は窃盗だと僕は思いますね。
上腕と前腕部が長く、海上での滑空巡航に適しつつ、獲物発見時には鋭い方向転換も可能らしいです。
あと、種小名からすると尾椎の長さに特徴があるんでしょうか?

鳥の趾は基本的に片側4本ずつで3本が前を向き第1趾が後ろを向く三前趾足が最もオーソドックスですが、その他にこのネッタイチョウのようにすべての趾が前を向く皆前趾足など複数のバリエーションがあります。とてもじゃないですがおぼえきれません。

潜水してエサを食べる鳥は肋骨が「く」の字に折れ曲がって胴体の後方まで包み込む形をしており、水圧から内臓を守っているとの事。でもそうじゃない鳥も結構「く」の字になってるように見えるやついるんですよね。

ペンギンの前肢はもう恐竜類じゃないだろこれ(恐竜類です)。

チドリ類は砂浜を走り回って小動物を襲うイメージですが、ブッポウソウのように滑空しながら虫を口を開けて虫を捕らえるそう。その生態が骨格に現れており、腕骨と前腕部が相対的に長く、空中での急旋回や長時間の滑空が得意らしいです。いろんなチドリがいるんですな。


普通、この骨は薄い板状らしいのですが、カモメ類では2つの穴が貫通し、軽量化に貢献しているそう。この部分は初列風切羽がつく部分ではありますが、強度が必要な部分ではないそうで、他のグループでも穴は無いまでも非常に薄いそうです。カモメ類はこのパーツに関しては、最も派生的な恐竜と言えそうです。

ミズナギドリやアホウドリのなかまは洋上の風速勾配を利用して滑空する、ソアリングと呼ばれる飛び方をするそうです。そういった鳥は前肢(翼)の骨が全体的に長く、後肢が短い傾向があるらしいです。

シギ亜目 Scolopaci とチドリ亜目 Charadrii は共にチドリ目 Charadriiformes であり、似た環境でしばしば共存していて行動も似ている為、まとめて「シギチ」と呼ばれますが、系統的にはシギ科はチドリ科よりもカモメ亜目 Lari に近縁だそうです。そんな系統的な差が骨にも現れているかもしれないので探して見てください(丸投げ)。

ハチドリ類 Trochilidae は世界最小の恐竜類です。体が小さいので、相対的に体に対する竜骨突起がでかいことでかいこと!ホバリングをするので、翼の高速回転に使う胸筋の付着部を稼ぐ必要性がある道理です。その一方で上腕骨は短くなっているのがわかりますでしょうか。
そしてこんな小さな小さな骨格標本を作れてしまう技術に脱帽です。

飛べる鳥の中で最重量級と言われ、体重を支えやすいようなのか、趾が短くなっているのがノガンの魅力です。
骨格から伝統的にはツル類 Gruidae に近縁とされてきた歴史があるそうで、湿地に適応したのがツル、乾燥地に適応したのがノガンと言われていましたが、流行りのDNA的にはツルとは遠縁でカッコウに近縁らしいです。生物なんもわからん。

オオバンは鳥の博物館が所在する我孫子市の鳥です。オオバンの骨の特徴は知ってますよ。膝(脛足根骨の近位前側)に注目すると大きな膝パッドがあるのがわかると思います。

これは筋肉の付着部の拡張の役割があり、クイナ類の共有派生形質なのですが、オオバンでは他のクイナ類と比べてこのパーツが大きく目立つのです。オオバンの足は弁足と呼ばれる、水かきとも違うヒレのついた趾になっており、水上をあしこぎで泳ぎます。水の抵抗を難なく蹴り進むため、陸生のクイナ類よりも大きな筋肉が要るのだと説明されます。

クイナ類は基盤的には飛翔性なのですが、島に定着した種は飛ばなくなる進化をする例がしばしば知られています。沖縄本当に生息する本種は日本ではその代表例で、竜骨突起や前肢といった飛ぶための構造が退化、足根中足骨が太く、骨盤が高くなるなど歩くのに適した骨になっているそうです。
余談も余談ですがヤンバルといえば、わたくしケラトプスユウタ、ジャングリアをボイコットしています。

セキセイインコの骨が世界でも有数にかわいいという半ば常識めいた感覚に触れたとき、この可愛さはいったいどこから生まれどこへ帰着するのかと哲学的な問いに身をよじらせた経験はきっと誰しもあるはずであり、しかもそれは骨にとどまらず標本全体から馥郁とした幸福の気配が立ちのぼって胸を締め付けるがゆえに、世界の宝として当然に問われるべき主題なのだそうです。小鳥ソムリエによると。
ちなみに、インコ目の多くで上腕骨の内部の中空部分の中で、どれだけの割合が空気で占められているか を示すパーセンテージ(ASP_i, Air Space Proportion of the internal cavity)が平均75%くらい(高度含気)である一方、セキセイインコは例外的に0%(完全に非含気)で、理由は不明らしいです(Burton et al. 2023)。うわすっげ!

羽毛に覆われた生体や剥製では絢爛な羽衣に目が行きがちですが、骨格にすると頭骨の巨大さに視線が奪われます。頑丈な下顎骨は種子を固定するのに適し、強靭な上顎で挟み込んで挫滅させます。上顎の付け根は薄く、ここでクチバシを下に曲げて強力に締め付けることができるそうです。恐い!!

恐竜の脚かと思ったら恐竜の脚でした。
ちなみにメスの方がオスより圧倒的に大きく、博物館に展示されているのはメスであることが多いらしいです。
***
感想:
まず関心したのが、展示は科や目といった分類枠を基準にしつつも、トピックごとに標本が横断的に配置されていたことです。例えば「飛翔」「潜水」「採食様式」など、機能やテーマで標本が並んでいました。これがとても良かったです。同じ分類群内での比較だけでなく、異なる系統間での収斂や差異が一目瞭然なわけです。これは単純に系統樹をなぞるだけでは見せられないものです。比較解剖の面白さを、我々にも直感的に伝える設計でした。
そしてなんと言っても、同一個体由来の「剥製 × 骨格」セットの「両取り」と呼ばれる贅沢。今回いちばん驚いたのがこれです。当然全てではないですが、多くの標本が同一個体由来の剥製と骨格標本のペア展示。これは本当にすごいことです。外形(軟組織込みの形態)と骨格を個体差ゼロの条件で比較できるわけです。通常の展示では「個体差」「年齢差」「性差」というノイズがつきまといます。両取りの標本に関してはその懸念がありません。だから理解がぱっちりクリアです。
そして何よりその技術的レベルの高さ!これですよ! 解体、処理、保存、組立。どなたか存じ上げませんがどれも神がかっています。平然と置かれていますが、裏側の仕事量を想像すると脱脱帽帽です。
次に狭さです。正直に言うと、広大な展示空間ではないです。それが良い。情報量が適切で展示によっては座ってじっくり観察できました。
巨大展で僕が陥る情報氾濫による時間切れも起きません。コンパクトだからこそ十全に伝わる展示意図があります。
展示内では、恐竜から鳥への進化の説明もきちんと用意されていました。ただ、これは当ブログでは何度か扱ってきた話題です。
むしろ僕にとって本展の魅力は、「鳥になった後」の骨の洗練と多様化を見せる点にあると感じました。
それから、国立科学博物館の『特別展 鳥』でサケビドリの前肢のトゲなど疑問に思っていた点があったのですが、今回の展示で、その疑問の一部が解消され、次の疑問につなげることができました。
展示を見に行くというのは単に知識を得る行為ではなく、自分の中の未整理な問いを更なる問いへと分岐する行為でもあるのだと改めて実感しました。
この恐竜展は、エンターテイメントを表に出すチャラチャラした展示会ではありません。かといって絶滅危惧や環境問題を問題提起する硬派な展示会でもないです。比較の妙、標本技術の高さ、コンパクトだからこそ一貫した展示意図。これらが、静かに積み重なっています。
手賀沼を歩き、鳥の博物館に入り、骨を拝む。それだけです。文句なしに満足して帰れる展示でした。

当ブログは旅ブログでもあるので遅いランチも紹介しますよ。
我孫子市内のラーメン屋『てらっちょ』の商品名は忘れましたが一番王道っぽいラーメン。こってり系です。おいしいです。

快晴でした。
また次の記事でお会いしましょう。
それじゃ👋
***
参考文献
- 川上和人・中村利和(2019)『鳥の骨格標本図鑑』BIRDER SPECIAL,文一総合出版,東京.
- Turner, A.H., Makovicky, P.J. & Norell, M.A. (2007) ‘Feather quill knobs in the dinosaur Velociraptor’, Science, 317(5845), p. 1721. doi:10.1126/science.1145076
- Alexander, G.D. et al. (1994) “A possible acoustic function for the casque structure in hornbills” Journal of Zoology, 232: 411–423.
- Burton, M. G. P., Benson, R. B. J. and Field, D. J. (2023) ‘Direct quantification of skeletal pneumaticity illuminates ecological drivers of a key avian trait’, Proceedings of the Royal Society B: Biological Sciences, 290(1995), 20230160. doi: 10.1098/rspb.2023.0160.
