

「予想通りになるなんて事ほどつまらない物はない( ・ω・)」山本聖士

ギガントラプトル Gigantoraptor は大型のカエナグナトゥス類 Caenagnatidae。一般的にはでかいオヴィラプトル類 Oviraptoridae のように思われる事があるが、オヴィラプトル類とは細部が違う。
多分、腕には羽が生えていたと思われるが、それがダチョウのような羽だったか、ただの繊維状だったのかはわからない。
オヴィラプトロサウリア Oviraptorosauria は通常、羽毛恐竜のように復元されるがカウディプテリクス Caudipteryx 以外の直接証拠はない。
オヴィラプトロサウリアはスカンソリオプテリクス類 Scansoriopterygidae に近縁。
羽毛恐竜は探すようになったら見つかるようになった。大概は水分を失ってボロボロになる。
ギガントラプトルは羽毛が生えていたとしても、確実に非対称の(前縁が狭く後縁が広い)羽毛ではあり得ない。飛ばなくなった鳥は二次的に左右対称の羽になっている。極端化するとキウイのような羽毛になる。
ギガントラプトルは走鳥にないようなプロトフェザーのような羽毛もあったかもしれない。
鳥は羽のあるところとないところがあり、長い羽によって羽がない部分を覆っている。
中生代は全体的には寒暖差がなく温暖だったので、小型種よりは羽毛みっしりではないはず。
緯度や標高によっては寒冷な気候だった地域もあったはずだが、ギガントラプトルは植生豊かな土地(温暖な土地)に住んでいたと思われるので、身体全体を覆う感じでは生えていなかっただろう。
脚は長いが、脛が大腿部より少し長い。これは大型二足歩行の恐竜には珍しい。大型二足歩行動物の脚は、遠位(つま先に近い方)ほど短くなるのが普通。小型の近縁種よりは短くはなっている。
尾が短いのに全長6mほどに達し、同じ長さの他の恐竜に比べて2倍ボリューミィ。
体高的にはキリンっぽい。
普段住んでいる環境と卵を産む環境は違うのが普通。特に大型のものは機動力があるので積極的に場所を選ぶはず。
ギガントラプトルの詳しい骨学的研究はまだ。らえらぷすさんの図ほども実際には見つかっていない。
マウントの胸骨はひどいアーティファクト。腸骨は背側が欠けている。
カエナグナトゥス Caenagnathus の既知の部分は、ギガントラプトルとの類似点が多い。
ギガントラプトル自体の骨は下顎以外は参考にできない。
短報で使われた下顎の図は実物と違う。
(日本人コレクターは最悪。売る側はヤクザ。)
カエナグナトゥス類はアークトメタターサリアン。第1趾が残っていてオルニトミムス類 Ornithomimidae と違う。前肢がしっかりしているのでティラノサウルス類 Tyrannosauridae とも違う。
オヴィラプトル類はアークトメタターサリアンではない。アークトメタターサルはオヴィラプトロサウリアの基本構造ではない。
ギガントラプトルはカエナグナトゥス類が知られるようになった頃にセンセーショナルに発表された。
下顎の先がオヴィラプトル類とカエナグナトゥス類はけっこう違う。開口部を途中で分けるような突起がカエナグナトゥス類にはない。肉つけたらわからないけどね!
先祖は恐らく樹上性で翼を持っていた。そうでないと尾が短くて前肢が長いことの説明がつかない。
鳥のような胸郭のつくりで、関節面が横を向いている。つまり羽ばたきができる。
胴と尾が極端に短い。オヴィラプトロサウリアも短いが、輪をかけて短い。
カウディプテリクスに類似性が見いだせる。
鳥は腸骨が前側に固定的になっているので、バランスを取るために跗蹠を伸ばすしかないから、跗蹠が例外的に長い。
アヴィミムス Avimimus は骨盤後方が極端に大きくなっている。
(中国の論文は本当に短報だけで実績を稼ぐのが多い。他のタクサとの違いを述べておしまい。そういうのは中国の恐竜ではよくある)
翼を日除けにしたかも。先祖は滑空などのために使ったと思われる。
体重は当初2tと見積もられていた。現在は数百kg、重くて800kgと推定される。
卵は複数の個体が一つの巣に産んだと思われる。一腹につき2個っぽい。卵のサイズは生物史上最大級(マクロエロンガトーリトゥス)。
抱卵した姿で発見されたキティパティ Citipati などは子育て恐竜の一つと言われるが、卵の保護と子供の保護は直結しないよね。
【頭】

カエナグナトゥス類全体でも頭骨は最も未知。アンズ Anzu の発見でそこそこわかってきたが、それも破片なのでな。
下顎がカエナグナトゥス類を特徴づける。オヴィラプトル類よりも低いクチバシを持っている。下顎の先が先細りのU字型。オヴィラプトル類の下顎の先は角ばっている。
ギガントラプトルの下顎はアンズのより背が高い。
ギガントラプトルの方が基盤的。
でもそれに見合った上顎の形をしていない。
アンズの復元頭骨(下図)は下顎尖らせすぎ。

レプトリンコス Leptorynchos の下顎は絶壁になっている。
顎の違いは食性の差。食べわけができるということは、白亜紀末まで植生が維持されていた証拠で、白亜紀末に多様性が損なわれていたという説は否定されている。
恐らく鉤状突起は上顎にはないので肉食ではないが、植物食にしては胴体のボリュームが小さい。
下顎の先の形状は重要で、少なくともこれに被さる形状の上顎を持っていたことが推定できる(でもオヴィラプトル類の上顎の先は角ばってないんですけどね)。
下顎は左右でくっついた状態で生まれてくる。少なくとも先端で扇状に広がる上顎ではない。
何を食べていたのかは謎だが、基本的に植物食。ダチョウは長い脚に合わせた長い首を使って低いところにある植物を食べる。
時期によっては動物質のものをメインに食っていた可能性もある。
オヴィラプトル類は恥骨が後ろにカーブしている。つまり腹のスペースを確保している。
カエナグナトゥス類は全体的にオヴィラプトル類より背が高い。
オヴィラプトル類は前上顎骨が高度に鼻骨と癒合している。カエナグナトゥス類は鼻骨と癒合しない(明確にトサカがあるのになぜなのか?)。
牙状突起が口の奥の方にある。舌と共に使うことになる構造(口蓋歯)。
口蓋歯は恐竜ではほぼ見られないので珍しい。口蓋歯自体は爬虫類にありふれている。鳥に近い恐竜にもそういうのが発達する因子を持っているってことよね。口蓋歯は爬虫類の中でも生える場所にバリエーションがある。歯は表皮構造ならどこでも生え得る。
オヴィラプトル類のクチバシを支える骨は前上顎骨だけ。カエナグナトゥス類のクチバシを支える骨は前上顎骨と上顎骨の二つ。
カエナグナトゥス類は目より前の部分が長く、下のクチバシの前方と対応しているように見える。
オヴィラプトロサウリアのトサカが高いやつは前頭骨からすでに高くなっている。エルミサウルス Elmisaurus のは低いので、トサカの後ろの方は高さがなかったかも。
目の後ろの穴が小さいのは頭骨の可動性と関係ある。
(一応妄想に根拠がないわけじゃないんですよ、という話がしたいよね)
鼻骨がなかなかに2次元的な形をしている。
意外と鼻孔がでかいのではないだろうか。
下顎後方は前後の可動性があると思われる。前方はそんなに可動しない。
涙骨は薄そうだが前方に張り出しの構造があるかな。
アンズの後眼窩骨の関節が緩いのは確か。
アンズでも鼻骨は未発見なのでカエナグナトゥス類のトサカはオヴィラプトル類のを参考にするしかねえ!
「博物館に展示されているのがすべて妥当な復元だと思ったら大間違い」
眼は真横を向くはず。眼のつき方からすると積極的な捕食動物ではない。
断片でも今まで出てなかった物が出ている。それだけで新たな事がわかる。
相対的な咬合力はオヴィラプトル類の方が強い。カエナグナトゥス類はシャベルのようにすくうような物っぽい形への定向性がある。
【首】
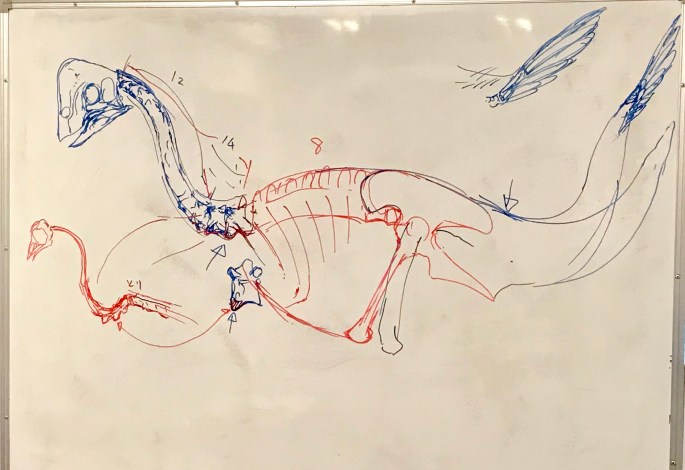
頸椎12個。頸椎化した胴椎が2〜3、胴椎7〜8。
アンズは頸椎自体が2つ増えており、非鳥類獣脚類ではすこぶる珍しい。
ギガントラプトルは高いところにある葉を食った可能性がある。
頸椎の背側の突起は小さく、筋肉は少なかった。
首は持ち上げ気味で後ろに引くのがデフォルトかな!
【胴体】
前方胴椎の首の付け根側の下側に突起がある。これは鳥類にも見られる。本来のS字構造の下に支えの構造がある。
普通の獣脚類より短くて丸っこい胴体。
アパトラプトル Apatoraptor の標本で肋軟骨の構造が残っていて、胸郭の高さ方向の厚みが明らかになった。
前から見ると烏口骨が相当前側にうねっている(ギガントラプトルのマウントは一応それを意識しているんだな)。
そんなに薄い胸郭じゃないが、デイノニクス Deinonychus と比べると薄い。
肋骨の左右幅が広いのは上側ではなく上下。丸っこいと言ってもティラノサウルス Tyrannosaurus とは違う。
肋骨は長時間植物を消化するので、常に膨らんだ状態と思われる。
腸骨はカエナグナトゥス類ではノミンギア Nomingia でしか完全な物は知られていない。オヴィラプトル類の恥骨は後方にカーブしているが、カエナグナトゥス類ではまっすぐなので大きな内臓を入れる余地がない。
【前肢】

腕はデイノニクスより低い位置になる。
この構造はカエナグナトゥス科内での差はないはず。
ギガントラプトルは相対的に手がでかい可能性はある。
腕のプロポーションはタニコラグレウス Tanycolagreus に似ているが、爪が違う。爪はテリジノサウルス類 Therizinosaurid に似ている。
爪は関節に対して大きく飛び出した構造で指を大きく動かせる。
へユアンニア Heyuannnia やキティパティは指の長さが揃っているが、キロステノテス Chirostenotes の指の長さは揃っていない。獣脚類は揃ってないのが普通だが、オヴィラプトロサウリアでは揃っているのが基本。第3指が長い恐竜を祖先に持っているのかもしれない。
手に羽毛を生やしたら外見では表現しづらい。
羽毛を剥ぐと手羽先に鉤爪が生えているような感じ。翼面積を稼ぎ、回内をするために橈尺骨が湾曲している。
飛ぶ鳥の翼をそのまま参考にできるが、羽の復元は要注意。上腕の近位少なくとも三分の一くらいは胴体に埋もれていたはず。
肩から手首にかけて張られた膜はアンキオルニス Anchiornis から知られている。生体では羽毛に覆われているので膜状には見えない。
ギガントの腕はよく発達しているように見えるが、飛翔性の祖先の名残に過ぎず、退化傾向にあると思われる。より派生的なアンズの方が小さいし。
地上性の二足歩行獣脚類の前肢は通常あまり機能的ではない。
【後肢】

大腿部に対して脛がかなり長く鳥類っぽい。中足骨が脛の半分くらいの長さ。オヴィラプトル類もそれに近い。
カエナグナトゥス類の後ろ足は癒合度がものすごく高いのが知られている。
ギガントラプトルではそうでもないけど足根骨を膨らまし気味にするのが望ましい。
足指は第3が長く、走行性を示唆する。
足首は膝より少し前に出る。その点はティラノに近い。
【尾】
尾は可動性が高くかなり上に跳ねあげられる構造。下側は血導弓が発達しているのであまり曲がらない。
尾端骨は多くの獣脚類が持っていた構造。
尾は太短いが大きな尾大腿骨筋が入っていたと思われる。地上性になるために厚みを増してバランサーとして機能していると思われる。
以上だばーろー。
